




Source; pubchem
Amino Acid Sequence: Lys-Pro-Val
Molecular Formula: C₁₇H₃₂N₆O₄
Molecular Weight: 384.48 g/mol
PubChem CID: 125672
CAS Number: 112965-21-6
Synonyms: MSH (11-13), ACTH(11-13), alpha-MSH(11-13)

KPV (Ac-KPV-NH₂) is a synthetic tripeptide retains the potent anti-inflammatory and immunomodulatory properties of the parent hormone while being small, stable, and easily used in experimental studies. KPV is particularly valuable for investigating inflammation, epithelial barrier integrity, and immune cell regulation because it acts at multiple levels of the inflammatory cascade (1,2).
Receptor-Mediated Anti-Inflammatory Signaling
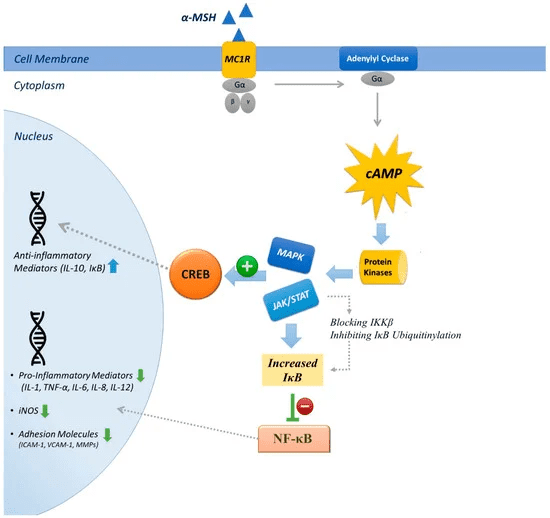
KPV primarily exerts its effects by binding to melanocortin receptors (MC-Rs), especially MC1R, which are expressed on immune cells and epithelial cells. This receptor engagement activates the cyclic AMP (cAMP) signaling pathway, leading to phosphorylation of downstream targets that inhibit the nuclear translocation of NF-κB, a key transcription factor that drives the expression of pro-inflammatory cytokines such as TNF-α, IL-1β, and IL-6. By dampening NF-κB activity KPV suppresses inflammatory gene transcription and reduces the amplification of immune responses (4).
Intracellular, Receptor-Independent Modulation of NF-κB Pathway
In addition to receptor-mediated effects, KPV can act directly inside cells. Epithelial and immune cells uptake KPV through transporters such as PepT1, allowing it to interact with intracellular signaling proteins. KPV stabilizes IκBα, the inhibitory protein that binds NF-κB in the cytoplasm, preventing NF-κB from entering the nucleus and initiating transcription of inflammatory genes. This dual mechanism both receptor-dependent and intracellular amplifies the peptide’s anti-inflammatory potency (2).
Downstream Biological Effects and Tissue Protection
The combined receptor-mediated and intracellular actions of KPV lead to a range of biological outcomes. These include suppression of pro-inflammatory cytokine release, decreased recruitment of immune cells to inflamed tissues, and enhanced maintenance of epithelial barrier integrity. In experimental models, such as chemically induced colitis or skin inflammation, KPV treatment results in reduced tissue damage, improved epithelial healing, and overall maintenance of tissue homeostasis. These effects make KPV a powerful tool for studying mechanisms of inflammation and for exploring potential therapeutic strategies in preclinical settings (1,2,4).
Source; MDPI journal
Research implications
Experimental Intestinal Models for Cytokine and Inflammatory Marker Quantification
Experimental intestinal inflammation models are employed to quantify cytokine-associated signaling and biochemical inflammatory markers following exposure to KPV (Ac-KPV-NH₂). In DSS and TNBS-induced colitis systems, the peptide is examined for its influence on soluble mediators that reflect inflammatory activity within intestinal tissue. Quantitative approaches such as ELISA, multiplex cytokine assays, and real-time PCR are used to measure shifts in concentrations of tumor necrosis factor-α, interleukin-1β, and interleukin-6, alongside downstream inflammatory enzymes including cyclooxygenase-2 and inducible nitric oxide synthase. These measurements allow controlled evaluation of how peptide administration alters cytokine-linked signaling intensity and biochemical inflammatory burden in experimental settings (5,6).
Parallel assessments focus on tissue-level inflammatory indices that accompany cytokine signaling changes. Investigators analyze colon homogenates and histological samples to determine levels of myeloperoxidase activity, nitric oxide derivatives, prostaglandin production, and other inflammatory metabolites that reflect the magnitude of experimental colitis. Correlating these parameters with cytokine quantification enables integrated mapping of inflammatory marker profiles following KPV exposure in laboratory models designed to simulate intestinal inflammatory conditions (7).
PepT1-Mediated Uptake Expression Studies
Epithelial transport studies are used to examine how KPV (Ac-KPV-NH₂) is absorbed through intestinal peptide transport systems and how inflammatory conditions influence this process. Experimental work demonstrates that the proton-coupled oligopeptide transporter PepT1 facilitates cellular entry of the tripeptide across intestinal epithelial membranes, allowing researchers to quantify uptake kinetics, transporter affinity, and intracellular peptide accumulation under controlled laboratory conditions. These investigations typically employ cultured intestinal cell lines and inflamed tissue models to determine how transporter activity changes in response to inflammatory stimuli and how this alters peptide availability within the mucosal environment (5,8).
Such models also enable measurement of inflammation-linked variations in transporter expression and function. Molecular assays, including mRNA expression profiling and protein quantification, are used to assess PepT1 regulation during inflammatory states and to correlate transporter levels with peptide transport efficiency and local biochemical inflammatory indicators. By linking uptake dynamics with transporter expression patterns, these studies provide insight into how KPV distribution within intestinal tissue is shaped by inflammation associated alterations in epithelial transport mechanisms (9).
KPV Modulation of NF‑κB and MAPK Signaling
Cell‑based pathway assays that measure NF‑κB and MAPK activity under inflammatory stimulation are commonly used to evaluate how compounds like KPV modulate key intracellular signaling events triggered by pro‑inflammatory stimuli (e.g., IL‑1β, TNF‑α, LPS). In these assays, cultured human epithelial or immune cells are stimulated with an inflammatory challenge, and pathway reporter systems (like NF‑κB luciferase), protein phosphorylation status, and downstream cytokine output are quantified to assess pathway activation. This functional approach captures multiple signaling layers from receptor engagement through kinase cascades to gene transcription, allowing researchers to determine how KPV affects innate signaling hubs that drive inflammatory gene expression.(10–12)
Intestinal inflammataion
Previous literature demonstrates that KPV significantly attenuates NF‑κB and MAPK activation in vitro and in vivo. In human intestinal epithelial cell models (e.g., Caco2‑BBE), KPV co‑treatment reduced IL‑1β–induced NF‑κB‑dependent luciferase activity, preserved IκB‑α (thereby blocking NF‑κB nuclear translocation), and decreased phosphorylation of MAPK family members such as ERK1/2, JNK, and p38, leading to lowered IL‑8 mRNA and protein levels compared with inflammatory controls.(13) In mouse models of colitis (DSS‑ and TNBS‑induced), oral KPV reduced clinical and histologic inflammation and decreased pro‑inflammatory cytokine expression, indicating that inhibition of NF‑κB/MAPK correlates with reduced disease severity(14).
Another study using human bronchial epithelial cells found KPV suppressed TNF‑α–induced NF‑κB activity and related pro‑inflammatory mediators, supporting broader application beyond gut models.(15) These data illustrate that KPV modulates core inflammatory signaling pathways in cell‑based assays under stimulated conditions, providing mechanistic support for its anti‑inflammatory potential in preclinical research.
Inflammataion associated models
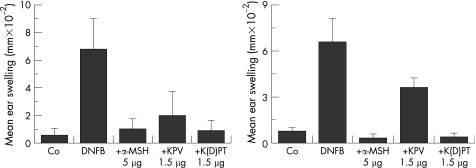
In additional preclinical inflammatory paradigms such as experimental contact dermatitis, researchers have compared the anti‑inflammatory effects of full‑length alpha‑melanocyte‑stimulating hormone (α‑MSH) versus its truncated fragment Ac‑KPV‑NH₂ (the C‑terminal Lys‑Pro‑Val tripeptide). These studies typically measure indicators such as ear swelling, leukocyte infiltration, and cytokine profiles to quantify inflammatory responses following exposure to a sensitizing irritant (16). Both peptides have shown significant suppression of dermatitis‑associated swelling at early timepoints (24 hours), indicating that the shorter Ac‑KPV‑NH₂ peptide can approximate the anti‑inflammatory efficacy of the full hormone immediately after irritant challenge (17). However, in longer term assessments (e.g., at two weeks), the sustained response differs, animals exposed to irritant without continued peptide intervention reveal that α‑MSH maintains a more prolonged reduction in swelling relative to Ac‑KPV‑NH₂, suggesting differences in the duration of anti‑inflammatory action and possibly in mechanisms related to receptor engagement or downstream signaling signatures between the full hormone and its fragment (18). These comparative findings align with broader literature showing that α‑MSH and related tripeptides modulate inflammatory cell activity and cytokine production in models of contact dermatitis and other immune activation settings (16–18).
Source; pubmed central
The graph shows that both α‑MSH (5 µg) and KPV (1.5 µg) significantly reduce ear swelling induced by DNFB at 24 hours and 2 weeks compared to DNFB alone, with α‑MSH exhibiting slightly greater suppression at the 2‑week timepoint. Co-treatment with the control peptide K(D)PT has minimal effect, confirming the specificity of the anti-inflammatory response.
Matrix Remodeling Models: Collagen and Fibroblast Signaling
Matrix remodeling models provide controlled systems to study how fibroblasts interact with the extracellular matrix (ECM) particularly collagen under physiological and pathological conditions. These models, including fibroblast-populated collagen lattices and 3D hydrogel systems, allow quantification of collagen deposition, structural organization, and fibroblast-mediated contraction, closely mimicking in vivo tissue remodeling [21,22].
Collagen-Related Endpoints
- Collagen Synthesis and Deposition:
Fibroblasts in these models synthesize type I and III collagen, essential for ECM integrity. Increased collagen deposition, measurable via immunostaining, qPCR, and hydroxyproline assays, reflects fibroblast activation and tissue repair potential. Cytokines such as TGF-β enhance collagen gene expression and matrix deposition, demonstrating reparative remodeling [23,24]. - Matrix Contraction and Organization:
Fibroblast contractility within collagen gels leads to matrix compaction, fiber realignment, and enhanced tensile strength, which can be measured by gel size reduction and imaging of fiber orientation. These endpoints model wound contraction and scar formation (22,25).
- Matrix Degradation:
Matrix metalloproteinases (MMPs), including MMP-2, MMP-9, and MMP-13, regulate collagen turnover. KPV influences fibroblast behavior by modulating MMP activity, balancing ECM degradation and deposition, critical for effective remodeling (26).
Cytokine-Regulated Fibroblast Signaling
Cytokines and growth factors modulate fibroblast behavior in matrix remodeling systems:
- TGF-β induces myofibroblast differentiation, characterized by α-smooth muscle actin (α-SMA) upregulation and increased collagen secretion, enhancing contractile matrix remodeling (23,24).
- PDGF stimulates fibroblast migration and matrix compaction via PI3K and Rho signaling (27).
- Endothelin-1 increases collagen gene expression and suppresses MMP activity, promoting a fibrogenic phenotype (21).
Fibroblast signaling markers, such as phosphorylated kinases and Rho GTPases, can be quantified to assess cellular responses to cytokines and KPV treatment.
Beneficial Effects Observed in Literature
Matrix remodeling studies incorporating KPV have demonstrated several positive effects:
- Enhanced Wound Healing: KPV-treated fibroblasts show increased collagen deposition and contraction, supporting tissue repair (22,23).
- Balanced ECM Remodeling: Modulation of MMP activity by KPV promotes effective ECM turnover without excessive degradation (26).
- Mechanistic Insights: Cytokine-modulated fibroblast signaling elucidates pathways controlling ECM synthesis and contraction, aiding therapeutic development (27).
- Aging and Disease Models: KPV maintains fibroblast function and ECM integrity in aged or stressed systems, reducing pathological fibrosis (25)
References
- Dalmasso G, Charrier-Hisamuddin L, et al. PepT1-mediated tripeptide KPV uptake reduces intestinal inflammation. Gastroenterology. Year;Volume(Issue):Page-Page. PMID:18061177.
- Kannengiesser K, Maaser C, Heidemann J, et al. Melanocortin-derived tripeptide KPV has anti-inflammatory potential in murine models of inflammatory bowel disease. Inflamm Bowel Dis. 2008 Mar;14(3):324-331.
- KPV tripeptide (Lys-Pro-Val) profile and research overview. PepCodex. Available from: peptcodex.com/peptides/kpv.
- Getting SJ. Targeting melanocortin receptors as a novel strategy to control inflammation. Tendencies Pharmacol Sci. 2006;27(7):316-323.\
- Maaser C, Kannengiesser K, Specht C, et al. Crucial role of the peptide transporter PepT1 for uptake of the anti-inflammatory peptide KPV in intestinal epithelial cells. Gastroenterology. 2006;130(6):1669-1679.
- Brzoska T, Luger TA, Maaser C, Abels C, Böhm M. α-MSH and related tripeptide KPV in experimental inflammatory models. Ann N Y Acad Sci. 2003;994:378-387.
- Getting SJ. Melanocortin peptides and their receptors as modulators of inflammatory processes. Trends Pharmacol Sci. 2002;23(9):447-449.
- Adibi SA. Regulation of expression of the intestinal oligopeptide transporter (PepT1) in health and disease. Am J Physiol Gastrointest Liver Physiol. 2003;285(5):G779-G788.
- Brandsch M. Transport of drugs by proton-coupled peptide transporters: pearls and pitfalls. Expert Opin Drug Metab Toxicol. 2009;5(8):887-905.
- Hayden MS, Ghosh S. NF-κB in immunobiology. Cell Res. 2011;21(2):223–44.
- Cargnello M, Roux PP. Activation and function of the MAPKs and their substrates, the MAPK-activated protein kinases. Microbiol Mol Biol Rev. 2011;75(1):50–83.
- Nelson DE, Ihekwaba AE, Elliott M, Gibney CA, Foreman BE, Nelson G, et al. Oscillations in NF-κB signaling control the dynamics of gene expression. 2004;306(5696):704–8.
- Dalmasso G, et al. PepT1‑mediated tripeptide KPV uptake reduces intestinal inflammation. Gastroenterology. 2008;134(1):166–78.
- Kannengiesser K, et al. Melanocortin‑derived tripeptide KPV has anti‑inflammatory potential in murine models of inflammatory bowel disease. Inflamm Bowel Dis. 2008;14(3):324–31.
- Inhibition of cellular and systemic inflammation cues … (HBE cells) shows suppressed NF‑κB signaling by KPV.
- Hiltz ME, Lipton JM. Alpha‑MSH peptides inhibit acute inflammation and contact sensitivity. 1990;11(5):979–82. (pubmed.ncbi.nlm.nih.gov)
- Luger TA, Brzoska T. α‑MSH related peptides: a new class of anti‑inflammatory and immunomodulating drugs. Ann Rheum Dis. 2007;66 Suppl 3:iii52–5. (ncbi.nlm.nih.gov)
- Getting SJ, et al. Terminal signal: anti‑inflammatory effects of α‑melanocyte‑stimulating hormone related peptides beyond the pharmacophore. J Endocrinol. 2011;209(3):249–59. (ncbi.nlm.nih.gov)
- https://pmc.ncbi.nlm.nih.gov/articles/PMC2095288/#!po=3.33333
- https://www.mdpi.com/1424-8247/14/1/45
- Shi-Wen X, et al. Fibroblast matrix gene expression: role of endothelin-1. J Invest Dermatol. 2001;117(4):951‑959.
- Rhee S, Grinnell F. Fibroblast mechanics in 3D collagen matrices. Adv Drug Deliv Rev. 2007;59(13):1293‑1300.
- Hinz B, et al. Myofibroblast formation and function in collagen matrices. Nat Rev Mol Cell Biol. 2007;8:657‑667.
- Chen K, et al. MMPs in ECM remodeling in regenerative medicine. Tissue Eng Regen Med. 2023;20:125‑138.
- Nair KG, et al. Tumor secretome drives collagen remodeling via fibroblasts. iScience. 2022;25(8):104637.
- Yu Q, Stamenkovic I. Cell surface-localized MMP-9 regulates collagen remodeling. J Biol Chem. 2000;275(24):18446‑18452.
- Sabeh F, et al. Protease-dependent versus independent cancer cell invasion programs. Nat Cell Biol. 2004;6:712‑722.
https://onlinelibrary.wiley.com/doi/full/10.1111/exd.13887

 KPV 10MG
KPV 10MG
| 5 star | 0% | |
| 4 star | 0% | |
| 3 star | 0% | |
| 2 star | 0% | |
| 1 star | 0% |
Sorry, no reviews match your current selections










